
客服QQ:
3518394516

邮箱地址:
support@z-peptide.com

电话:
13770887460
3518394516
support@z-peptide.com
13770887460


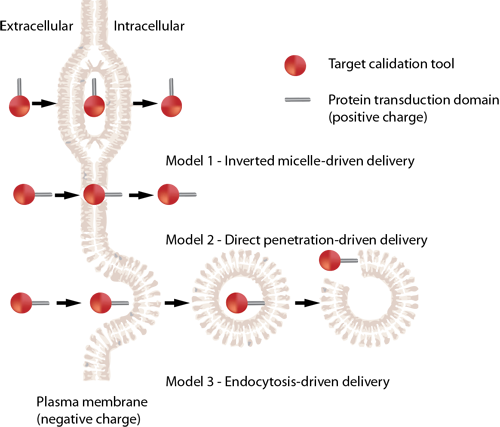
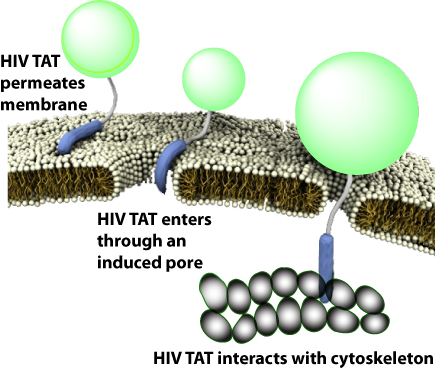
| 名称 | 来源 | 序列 |
| Tat family | ||
| Tat (48-60) | HIV-1 protein | GRKKRRQRRRPPQQ |
| Oligoarginine | Tat derivative | Rn |
| Penetralia family | ||
| p-Antp | Antermapedia homeodomain | RQIKIWFQNRRMKWKK |
| plsl | Igl-1 homeodomain | RVIRVWFQNKRCKDKK |
| Chimeric CPPs | ||
| Transportan | Galanin-mastoparan | GWTLNSAGYLLGKINLKALAALAKKIL |
| MPG peptides | ||
| P-beta | gp41-SV40 | GALFLGFLGAAGSTMGAWSQPKKKRKV |
| P-alpha | gp41-SV40 | GALFLAFLAAALSLMGLWSQPKKKRRV |
| Pep-1 | Trp-rich motif-SV40 | KETWWETWWTEWSQPKKKRRV |
2020-2021 正肽生化科技(南京)有限公司 版权所有地址:南京市江北新区智达路6号智城园区2号楼720-72室 苏ICP备2020050781号-1 XML地图